It’s full springtime, my favorite season.
Spring is full of metaphors (growth, new beginnings, resurrection) and always feels like a minor miracle to me. I love spring for the dynamics, the reminders of time passing, and the way it highlights plant diversity. Early spring is stick season’s reflection, spread out, lasting longer than I expect. The signs of an upcoming spring are hard to come by in winter - most trees are bare and look the same in their bareness. Spring brings out their differences, makes them obvious in ways that are hidden in both winter (all branches) and later in the summer (all leaves).
I grew up in western Kansas, with relatively few trees - they were mostly along creeks, at homesteads, or in the cities. On our farm we had many, but you could probably go between a quarter to a full mile in any particular direction before you found another tree. Now I live in New England, which, comparatively, is completely covered in trees. They are throughout our cities, covering the hills, and every “natural” untouched space has them. Amazingly, my town has mapped every public tree in its city limits. This doesn’t even count trees on private property!
Every (public) tree in Somerville, MA.
I’ve now lived here long enough to recognize the most common trees and note the sequence of all of their blooms every spring: magnolia, maple, cherry, Callery pear, dogwood, lilacs, oaks, black locust and onward. I’ve come to especially anticipate and love the season of cherry (sakura) blossoms. Of course, I’m not alone in loving cherry blossoms. Especially in Japan, their blooming is occasion for celebration (Hanami (花見), flower viewing). The Japanese “cherry blossom front” is tracked every year.
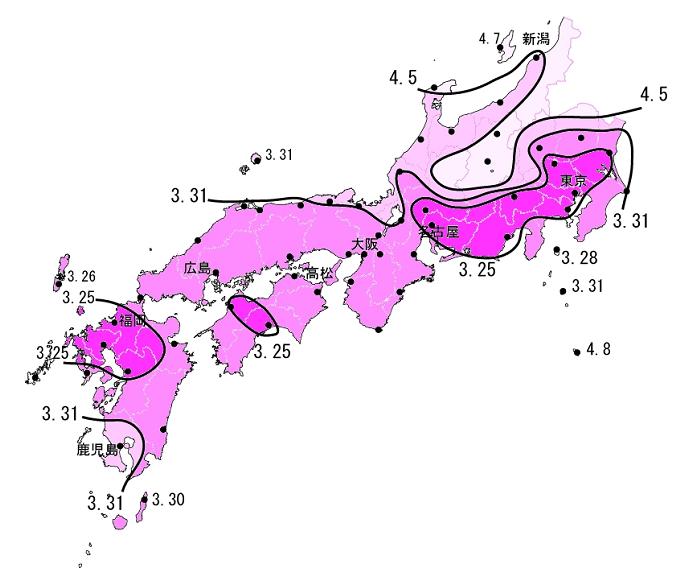
The cherry blossom front between Kyushu and Kanto, 2007. The numbers are dates, and the shading (dark to light) represents bloom times (earliest to latest).
I recently flipped through a lovely children’s book on display at Porter Sq Books, “When Cherry Blossoms Fall” by Katrina Goldsaito (author) and Yas Imamura (illustrator). It had a beautiful quote to the effect of “when you see a sakura flower in full bloom, it already feels like it’s behind you.” They evoke “mono no aware” (物の哀れ), or the bittersweet feeling of impermanence, which in fact is always simmering under the surface of things.
So, what controls the timing of blooming? Why nothing, then seemingly all at once? Why is the sequence of species so regular? What makes the world wake up from its winter every year?
In general, growth in plants is controlled by gene expression: which genes are being turned into proteins at any given moment. When it comes to blooming, the major causal environmental signals are temperature and photoperiod (day length). For temperate trees and many spring-flowering plants, the plant must remember that winter happened, but it must also notice that spring is happening now. It must also deal with day-to-day variability, so that it doesn’t overreact to one warm day in January or a hard cold snap in March. Coordinating all of this is important evolutionarily: bloom synchronization maximizes the chances of reproduction. The two timescales involved are read out by the plant in two different ways: the passing of winter is driven by a cold-sensing epigenetic switch that primes the whole plant for spring, and daily warm/cold swings are tracked by temperature-dependent transcript splicing and protein degradation dynamics. Together, these mechanisms act as a dual-lock system that must both be opened for the plant to produce flowers (1, 2).
To explore these ideas I made a small interactive model of this process. It is not intended to be a real cherry blossom forecaster and is far from capturing everything in precise detail (it is complicated! (2, 3, 4)), but I learned a lot putting it together. Honestly, I’m vastly oversimplifying so much science here, but the shape feels right and it was satisfying and thought-provoking for me (a non-botanist).
Weather as an input
The model starts with a daily temperature series, . For the synthetic figures here, that means an annual seasonal rise and fall plus day-to-day weather variability. Each day gets a single number: the daily mean temperature in Celsius.
where is the mean annual temperature, is the seasonal swing, sets the warmest part of the seasonal cycle near late July, and is autocorrelated day-to-day variability in the daily mean. Day 0 is July 20, so the coldest part of winter is in the middle of the trace. Below I set these to roughly map to the climate of New England and Japan.
Plants sense temperature in a variety of ways, including RNA structure, chromatin state, transcriptional regulators, cell membrane rigidity and protein stability (5, 6, 7). I’ve massively simplified this into two signals with smooth, dimensionless response functions:
and
where , and is a sharpness factor. These are essentially opposite logistic curves: is high below the chilling threshold and low when temperatures are too warm, and is the opposite, rising toward one as temperatures become spring-like.


The model
Our model has four state variables:
- : Polycomb silencing at a floral repressor locus. Polycomb-group proteins are a family of protein complexes that remodel chromatin, physically preventing gene expression from happening.
- : FLC/DAM-like expression. These are the genes that are epigenetically silenced. They induce dormancy and repress flowering, so when they are high they act like molecular brake on flowering. They’re integrators of many environmental signals.
- : SVP-like expression. This is another repressor of flowering, but its regulation is reversible and thus it can track current conditions.
- : florigen expression. This is a plant hormone protein that is produced in the leaves and acts as a universal regulator of shoot growth, including flowering.
The equations are:
Bloom happens when crosses a threshold, .

: winter memory
The first equation is an epigenetic switch:
The term writes Polycomb marks during effective cold. The term (especially the squared term) is the important nonlinear part: existing silencing helps recruit or stabilize more silencing. The term prevents from growing beyond 1, and is passive turnover.
This is meant to capture the intuition of vernalization. In Arabidopsis, prolonged cold represses the floral repressor FLC (FLOWERING LOCUS C) through Polycomb-associated H3K27me3 chromatin marks; that repression can be maintained after return to warmth. Reviews of the FLC system describe it as a beautiful example of environmental perception and epigenetic memory (3, 8).
: the floral repressor
The second equation translates chromatin memory into repressor protein:
The first term, , says the repressor is made when it is absent. The second term, , says Polycomb silencing pushes it down. When is low, tends to stay high. When flips high, falls and the first lock opens. In the annual Arabidopsis, this is FLC. In perennials like cherry trees, there is more to regulate, but homologs exist and the general principle of epigenetic silencing of a repressor holds (4, 9).
: the reversible warmth gate
The third equation is different:
recovers toward 1 when conditions are cold or neutral. Warmth pushes it down. This makes a current-weather gate rather than a winter-memory gate because there is no feedback loop to stabilize one state or the other.
The SVP (SHORT VEGETATIVE PHASE) genes are a useful biological analog. In Arabidopsis, SVP helps mediate ambient-temperature flowering responses and represses FT by binding regulatory motifs near the FT locus (10). Recent work also supports temperature-dependent SVP degradation through proteolysis (11). In trees, SVP-like DAM genes (DORMANCY-ASSOCIATED MADS-BOX) have expanded in woody plants, allowing for diversity in seasonal expression patterns and activity (4, 12). For this toy model, I collapse that biology into one reversible warmth-sensitive variable. Once it gets warm enough, DAM genes are degraded or down-regulated, which results in the expression of the genes they control.
Unlike the epigenetic regulation of , SVP/DAM is rapidly reversible. A warm spell in January can lower for a moment, but if the weather turns cold again, recovers. With appropriate temporal dynamics, SVP expression can sense and act as a continuous “low-pass filter” of warmth, rather than a binary on/off.
: the double-lock output
The final equation is the output:
FT (FLOWERING LOCUS T), or florigen, is produced only when both repressors are low. If is high, winter memory has not opened. If is high, present conditions are not warm enough. Only when and are low at the same time does florigen accumulate. The term lets the signal decay or dilute away.
FT-like proteins are mobile flowering signals made in leaves and transported to the shoot apex, where they help trigger the floral transition (1). In this model, bloom is the moment crosses a threshold, .
Why winter has to last
A switch-like memory is useful because weather is noisy. Fall has cold nights; winter has warm spells. Plants need to distinguish a cold snap from a season.
In the model, a brief cold pulse is not enough to push Polycomb silencing over the threshold. The marks decay back toward zero, the floral repressor stays high, and FT does not accumulate. Sustained cold crosses the threshold, after which the memory stays high even when temperatures rise (13).
We can explore this with a controlled lab-like protocol. Each run starts at a neutral holding temperature: too warm to count as effective chilling, but too cool to open the warmth gate very much. Then we can insert a controlled cold block, varying only its duration. After that, every run receives the same spring ramp. Once the memory switch flips, it is mostly permissive: the runs that clear the winter threshold bloom together because spring warmth is held identical.

In simulated lab conditions, cold duration can affect switch timing.
Climate change and bloom timing
The peak flowering day in Kyoto has actually been tracked since the 9th century, first through court diaries and other historical records, and later through modern observation. The resulting time series is one of the most remarkable biological climate records we have: a record of spring stretching across more than a thousand years (14, 15).
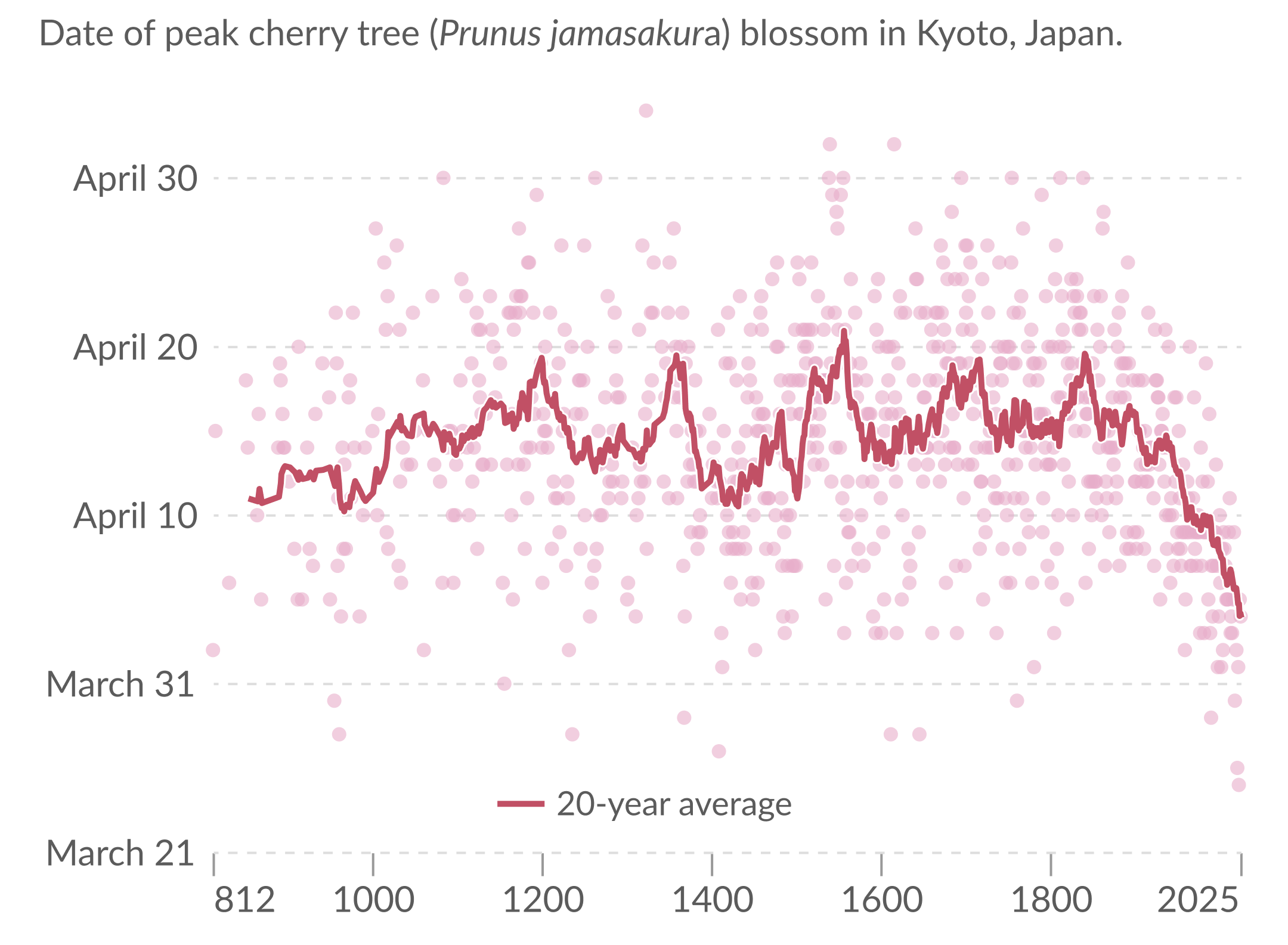
Our World in Data visualization of peak cherry blossom dates in Kyoto, using data from Yasuyuki Aono.
Warmer winters can reduce chilling and delay, weaken, or desynchronize the memory step. Warmer springs can also induce the warmth gate to open earlier. The balance depends on species, local climate, and the parameter regime. In the synthetic run below, spring warmth dominates and bloom advances with warming. Other choices can make chilling limitation show up more strongly (16, 17).

Warmer temperatures shift flowering earlier in the year.
| Temperature offset | Bloom shift vs. baseline |
|---|---|
| -2 C | 5 days later |
| +0 C | baseline |
| +2 C | 7 days earlier |
| +4 C | 27 days earlier |
Even in this toy version, the response is not subtle: a two-degree warming shifts bloom about a week earlier, and a four-degree warming shifts it nearly a month earlier. The influence of increasing average temperatures due to climate change is striking. Here is hoping it is not a sign of larger impermanence.
A sequence of species
Our toy model also suggests a partial answer to the regular spring sequence. Plants can regulate their bloom timing in a variety of ways. A species can differ in how quickly winter memory accumulates, how sensitive the warmth gate is, how fast FT is produced, or how much FT is required before bloom. Here I run the same synthetic weather against four parameter sets, varying the cold-memory rate, the SVP warmth-gate dynamics, FT production, and the bloom threshold. In this toy comparison, the winter switch has already flipped for all species. The sequence is mostly coming from the second half of the system, driven by daily temperature dynamics (2, 18).

Our model can reproduce the sequential bloom timing across species.
This is one of evolution’s great tricks: keep reusing a control architecture, but tune the knobs. Gene duplication and divergence can adjust the same kind of molecular systems into different seasonal behaviors.
All at Once
Spring is beautiful partly because it is brief, but also because of the depth and breadth of change it triggers. In addition to their visual beauty, flowers are the result of many interwoven and complex biochemical systems. In addition to form, their varied reactions to the seasons are another kind of biological diversity.



Yoshino blossoms at the Arnold Arboretum, spring 2026.
References
- Andrés, F. and Coupland, G. (2012). The genetic basis of flowering responses to seasonal cues. Nature Reviews Genetics.
- Lloret, A. and Badenes, M. L. (2023). Molecular mechanisms of flowering phenology in trees. Forestry Research.
- Whittaker, C. and Dean, C. (2017). The FLC locus: a platform for discoveries in epigenetics and adaptation. Annual Review of Cell and Developmental Biology.
- Falavigna, V. da S. et al. (2019). I want to (bud) break free: the potential role of DAM and SVP-like genes in regulating dormancy cycle in temperate fruit trees. Frontiers in Plant Science.
- Quint, M. et al. (2016). Molecular and genetic control of plant thermomorphogenesis. Nature Plants.
- Kerbler, S. M. and Wigge, P. A. (2023). Temperature sensing in plants. Annual Review of Plant Biology.
- Vu, L. D. et al. (2019). Feeling the heat: searching for plant thermosensors. Trends in Plant Science.
- Berry, S. and Dean, C. (2015). Environmental perception and epigenetic memory: mechanistic insight through FLC. The Plant Journal.
- Leida, C. et al. (2012). Histone modifications and expression of DAM6 gene in peach are modulated during bud dormancy release in a cultivar-dependent manner. New Phytologist.
- Lee, J. H. et al. (2007). Role of SVP in the control of flowering time by ambient temperature in Arabidopsis. Genes & Development.
- Lee, J. H. et al. (2025). Temperature-dependent SVP degradation through proteolysis. Nature Plants.
- Vimont, N. et al. (2020). Dormancy-associated MADS-box genes influence chilling requirement of sweet cherries and co-regulate flower development with SOC1 gene. Plants.
- Song, J., Irwin, J. and Dean, C. (2013). Remembering the prolonged cold of winter. Current Biology.
- Aono, Y. and Kazui, K. (2008). Phenological data series of cherry tree flowering in Kyoto, Japan, and its application to reconstruction of springtime temperatures since the 9th century. International Journal of Climatology.
- Aono, Y. and Saito, S. (2010). Clarifying springtime temperature reconstructions of the medieval period by gap-filling the cherry blossom phenological data series at Kyoto, Japan. International Journal of Biometeorology.
- Dai, J. et al. (2017). Timing of cherry tree blooming: contrasting effects of rising winter low temperatures and early spring temperatures. Agricultural and Forest Meteorology.
- Luedeling, E. et al. (2013). Identification of chilling and heat requirements of cherry trees — a statistical approach. International Journal of Biometeorology.
- Chuine, I. (2000). A unified model for budburst of trees. Journal of Theoretical Biology.